In Part 1 of our ‘Cell Structure and Function’ series we learnt about basic cell structure and the organelles. We now move on to the cell membrane and membrane transport.
The cell membrane, also called the plasma membrane, physically separates the inside of the cell (the intracellular space) from the outside environment (the extracellular space). It defines the borders of the cell and surrounds and protects the cytoplasm and cell contents. The cell membrane also allows the cell to interact with its environment in a controlled manner.
Membrane structure
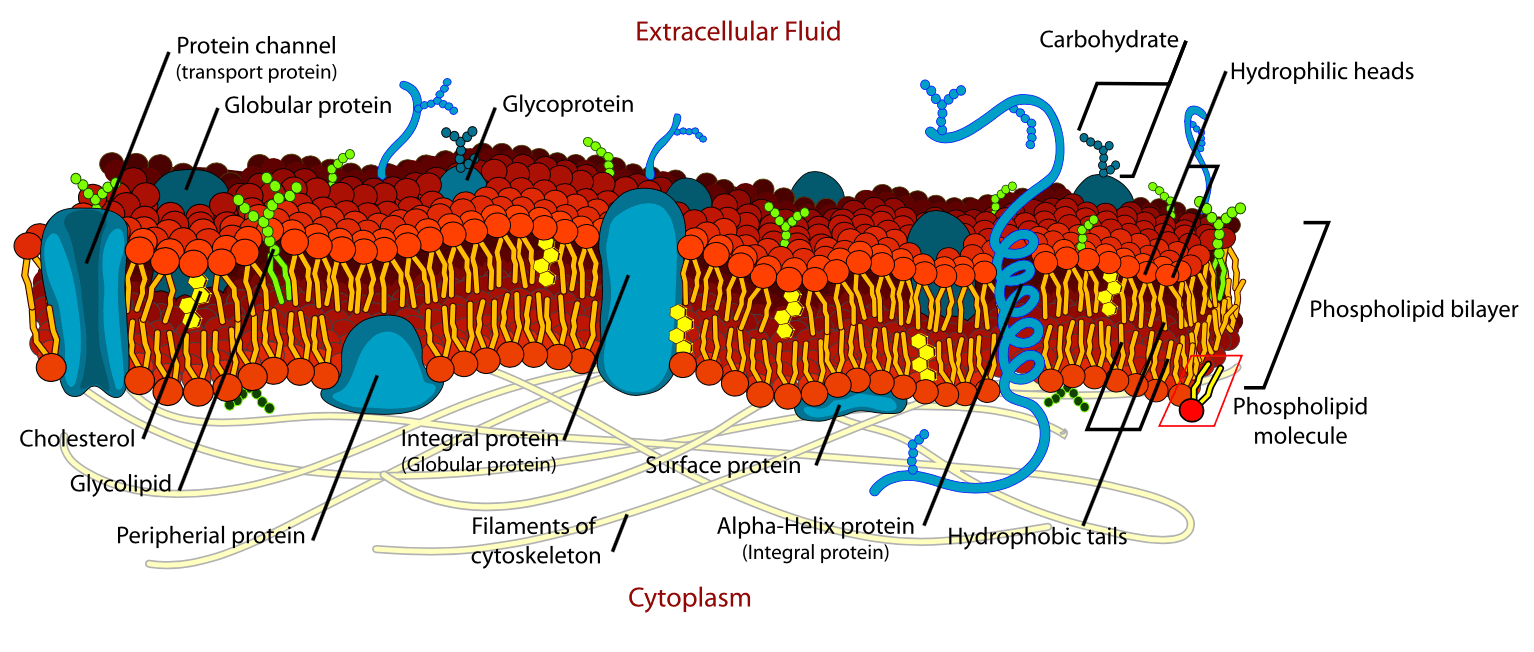
The cell membrane is composed of a double layer of phospholipids, which is referred to as a lipid bilayer. The phospholipids have a ‘water-loving’ (hydrophilic) head and a ‘water-fearing’ (hydrophobic) tail. The hydrophobic head is polar (charged) and can, therefore, dissolve in water. Conversely, the hydrophobic tail is non-polar (non-charged) and cannot dissolve in water.
In an aqueous environment, the polar heads attempt to form hydrogen bonds with the water, whereas the non-polar tails try to avoid the water. This results in the hydrophilic heads pointing away from the water and the hydrophobic tails point towards each another to ‘protect’ themselves from the water. The lipid bilayer, therefore, forms spontaneously due to the properties of the phospholipid molecules
Lipid molecules can diffuse freely within each layer, and the membrane is a fluid structure, with the lipid molecules continuously moving each other from side to side.
The structure of the lipid bilayer allows small, uncharged molecules like oxygen and carbon dioxide, as well as hydrophobic substances such as lipids, to move across the cell membrane by simple passive diffusion, following their concentration gradient.
In contrast, large polar molecules—such as glucose and amino acids—cannot pass through the membrane unaided and require the help of membrane proteins. Similarly, charged ions like sodium and potassium cannot cross the membrane on their own due to their electrical charge.
Membrane proteins
The lipid bilayer forms the basis of the cell membrane, but it is peppered throughout with various proteins.
Structural proteins, as their name suggests, provide support to the membrane. These are associated with the inner lipid layer and create a flexible support scaffold for the cell called a cytoskeleton. An example of a structural protein is spectrin.
Membrane proteins are proteins that span the membrane and have hydrophilic and hydrophobic regions that allow them to fit into the cell membrane. These membrane proteins act as carrier proteins that enable the transmembrane movement of the large polar molecules that cannot pass through by diffusion.
Glycoproteins are a type of integral membrane protein that have attached carbohydrate molecules that extend into the extracellular matrix. The attached carbohydrate tags on glycoproteins aid in cell recognition.
Glycolipids are a type of membrane protein that have carbohydrate chains attached to phospholipids on the outside surface of the membrane. These act as recognition sites for specific chemicals and are essential in cell-to-cell attachment to form tissues.
GTP-binding proteins (G-proteins) are a type of intracellular membrane protein that is associated with cell signalling. They are enzymes that cleave guanosine triphosphate (GTP) to guanosine diphosphate (GDP) to activate or inhibit other membrane-bound signalling enzymes, such as adenylyl cyclase, which generates cyclic adenosine monophosphate (cAMP) from ATP. cAMP activates protein kinase enzymes within the cell to instigate changes in cell function via the phosphorylation of intracellular proteins.
G-protein-coupled receptors (GPCRs) are a type of transmembrane protein that penetrates the entire thickness of the lipid bilayer. GPCRs detect and bind signal molecules, which in turn activates one of the G-proteins to initiate a biological cascade which results in altered cellular activity. GPCRs mediate most of our physiological responses to neurotransmitters, hormones and environmental stimulants and also have great potential as therapeutic targets for a broad spectrum of disease.
A diagrammatic representation of the structure of the cell membrane is shown below:
Membrane transport proteins
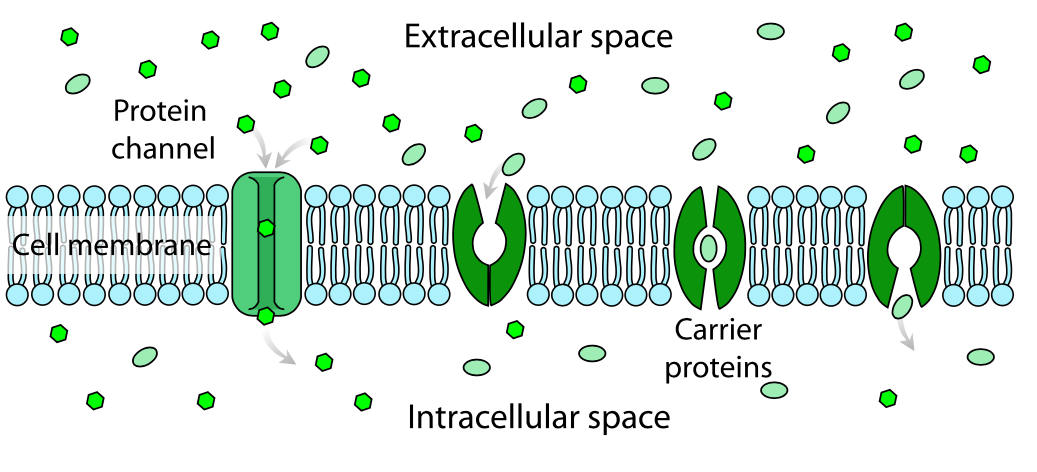
There are two main classes of membrane transport proteins; carrier proteins (also known as transporter proteins) and channels. Carriers are not open to both the extracellular and intracellular environments at the same time. Either the inner gate or outer gate is open, not both.
There are three main types of carrier protein:
- Uniporters: these move a single type of molecule in one direction across a membrane only.
- Symporters: these move several different molecules in one direction across a membrane only.
- Antiporters: these can move different molecules in opposite directions.
Molecules move through membrane carriers by facilitated diffusion, which is the movement of molecules down a chemical concentration gradient. No energy is required for this to occur. Facilitated diffusion differs from passive diffusion in that the transported molecules do not dissolve in the lipid bilayer. An example of carrier proteins is the GLUT glucose transporter proteins.
Unlike carrier proteins, channel proteins can open simultaneously to both the inside and outside of the cell, forming a continuous pore through which molecules can diffuse freely. Among the best-known examples are ion channels, which enable the selective passage of ions across the cell membrane. These channels are highly specific, allowing only ions of the correct size and charge to pass through their narrow pores.
Ion channels are regulated by “gates” that open temporarily in response to specific signals. Ligand-gated channels open when neurotransmitters or other signalling molecules bind to them, while voltage-gated channels respond to changes in the electrical potential across the membrane. Transport through these channels is extremely fast—around a thousand times quicker than transport by carrier proteins.
A diagrammatic representation of facilitated diffusion across a cell membrane is shown below. An ion channel is shown on the left, and a carrier protein on the right:
Molecules can also move across the cell membrane against their concentration gradient—from an area of low concentration to high concentration—through a process that requires energy input.
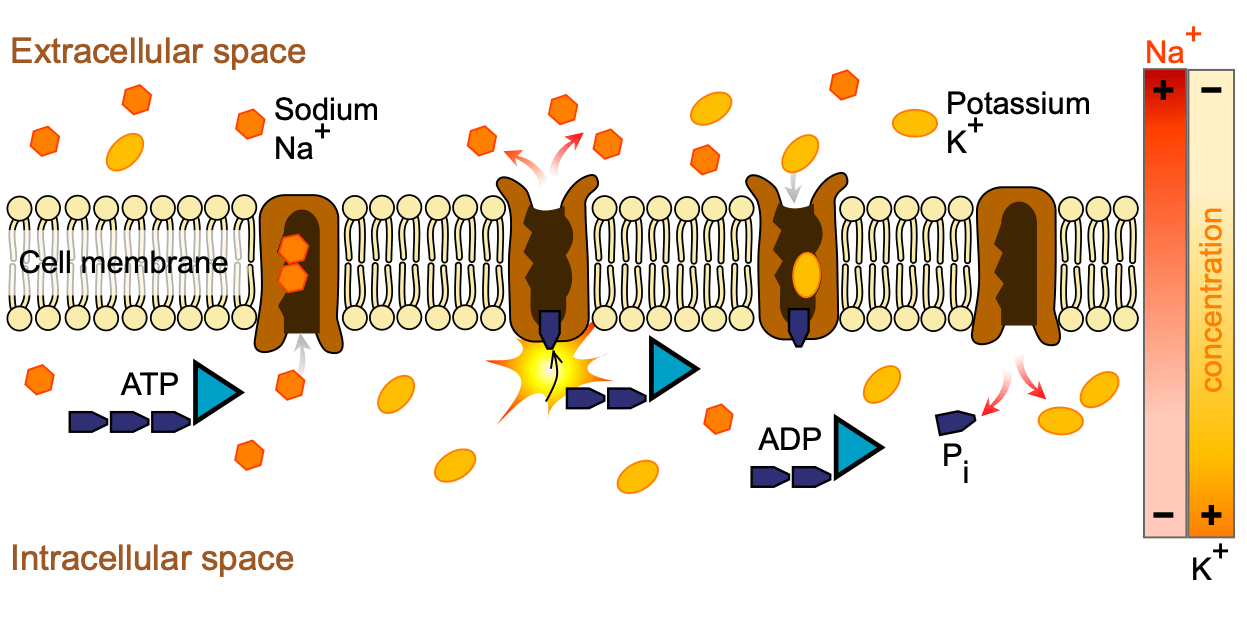
When this energy comes directly from a chemical source like ATP, the process is known as primary active transport. In secondary active transport, movement relies on an existing electrochemical gradient rather than directly using the cell’s own energy. In both cases, transport is selective and carried out by carrier proteins, with molecules moving either into or out of the cell.
A key example of primary active transport is the Na⁺/K⁺ ATPase pump. Located in the outer plasma membrane, this pump uses one molecule of ATP to move three sodium ions (Na⁺) out of the cell and two potassium ions (K⁺) in, both against their concentration gradients. This action helps maintain a high concentration of sodium outside the cell and a high concentration of potassium inside.
A diagrammatic representation of the sodium pump is shown below:
Header image used on licence from Shutterstock
Thank you to the joint editorial team of www.mrcemexamprep.net for this article.




